

Principales complicaciones logísticas en los arribos de uva de mesa a EEUU esta temporada
25/05/2022
Acuerdan avanzar en Systems Approach para las exportaciones de uva de mesa chilena a EEUU
09/06/2022
FUENTE : REVISTA MUNDOAGRO
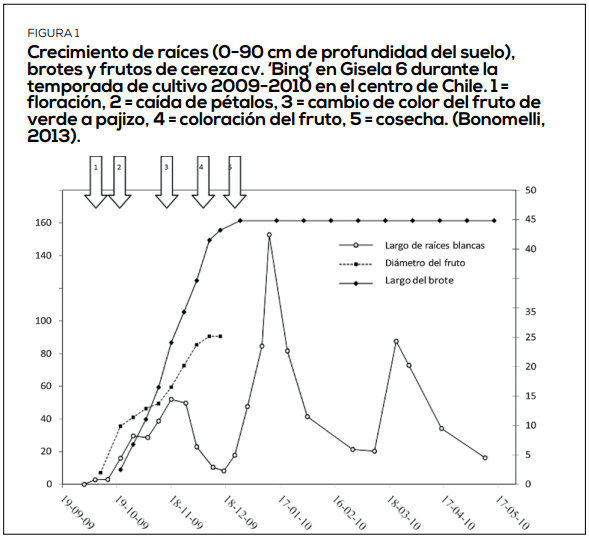
La inducción floral define el número y calidad de flores para la temporada siguiente, pero el estrés térmico e hídrico es algo que la afecta significativamente. En el cerezo, el proceso de inducción comienza unos 75 días después de plena flor (DDPF), lo que coincide con la reducción del fotoperiodo (20-25 de diciembre), cese del desarrollo anual de brotes y el comienzo de la inducción para la temporada siguiente y posterior comienza el periodo dormancia fisiológica. Estos eventos nos permiten entender la alternancia en la producción y el efecto de los estrés abióticos. Ver Figura 1.
En 2012 Nutriprove S.A. realizó ensayos pioneros de uso de preventores de estrés térmico e hídrico (PETH) en época de inducción floral (postcosecha) con la finalidad de reducir frutos dobles, reducir alternancia (estrés abiótico en época de inducción y diferenciación floral) y mejorar la producción de próxima temporada (vía aumento de las reservas nutricionales) y duración del follaje de postcosecha (para lograr mayor fotosíntesis y reservas).
El uso de PETH ya es considerada un estándar en la industria del cerezo en periodo de inducción floral en postcosecha ante estrés abióticos. (Fotos 1, 2, 3 y 4 de aplicaciones postcosecha y duración del follaje activo).

Estudios demostraron que temperaturas superiores a 20°C (desde un mes antes de floración) producen flores más pequeñas, con ovarios y óvulos más chicos, con sacos embrionarios débiles que degeneran rápido post floración (Beppu, 1997). Por esta razón es muy importante la prevención de estrés térmico un mes previo a floración hasta cuaja y posterior crecimiento de fruto para asegurar calidad de fruta.

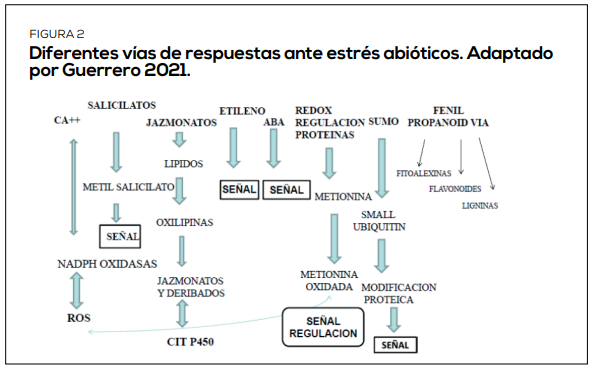
Preventores de estrés abiótico (PETH) de cuarta generación, considerados barreras físicas más fisiológicas y su mezcla con bioestimulantes como algas o aminoácidos es recomendable. La mezcla de PETH más bioestimulantes genera un efecto de baja de temperatura entre 3 a 5 grados en el follaje, asociado a una respuesta fisiológica de transformación de lípidos y otros compuestos como respuesta a estrés para hacer frente a las especies reactivas de oxigeno responsables de los síntomas de estrés (Figura 2). Además, es importante considerar de la mantención de la fotosíntesis, que ha sido medida en el caso de avellanos europeos (a modo de ejemplo) en las épocas de mayor temperatura y estrés.
RESERVAS-ESTRÉS
El crecimiento inicial del árbol depende de las reservas de la planta. Se ha demostrado que el desarrollo floral, la primera fase de crecimiento de los frutos y el desarrollo inicial de brotes y hojas son dependientes de las reservas acumuladas durante la temporada anterior (Loescher et al., 1990; Flore y Lane, 1999).
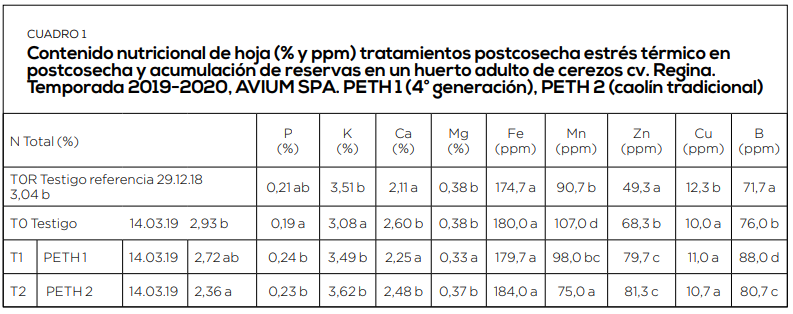
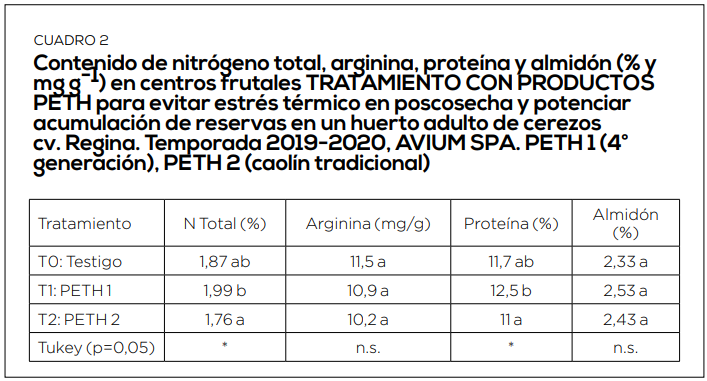
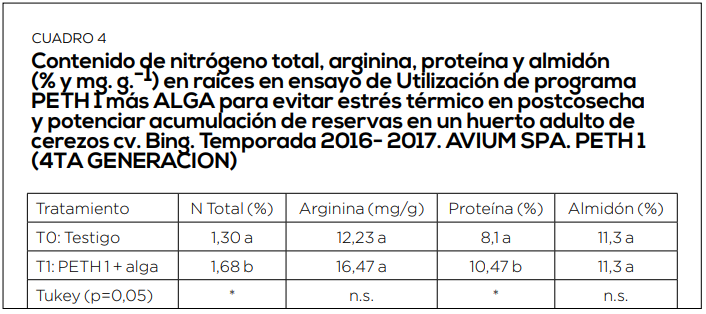
El periodo de postcosecha es el momento más importante de acumulación de reservas. Por ello es muy importante mantener activa la fotosíntesis hasta caída de hojas, el mayor tiempo y con la mayor eficiencia posible. Con ello, incrementamos en los niveles de arginina en raíces, los contenidos de almidón en dardos y estructuras florales. Y se reduce, de esta forma, la alternancia inducida por estrés abiótico. (Cuadro 3, 4, 5 y 6).
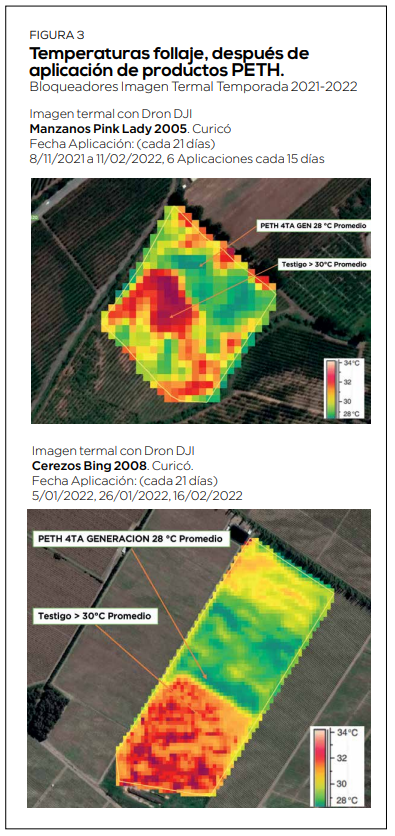
Varios de los procesos fisiológicos parten a los 75 días después de plena flor e influyen drásticamente sobre la viabilidad de floración y reservas (Figura 1, Bonomelli, 2013). Cuando las temperaturas superan los 25°C, se producen anomalías en el desarrollo de la yema, incidencia de espolones, suturas y frutos dobles. Enfriar el árbol desde fines de noviembre (incluso un mes antes de floración), reduce la incidencia de daños fisiológicos en próxima estación, mejora la calidad de la cuaja y los calibres en el año en curso. Nuestra preocupación entonces parte en la floración y se extiende a la postcosecha, con el objetivo de reducción del estrés abiótico térmico a través del uso de productos PETH (Figura 3).
TRANSPIRACION-CALCIO
El flujo xilemático de calcio y la transpiración de la fruta están relacionados (Brüggenwirth et al. 2016). La transpiración importa Ca a la fruta, pero el efecto es más débil en la etapa III de desarrollo del fruto y también en la madurez (Hirschi 2004; Park et al. 2005).
Existe una relación entre transpiración acumulada de frutos y la concentración de Ca del fruto de cerezos. Frutos con baja transpiración alcanzarán concentraciones de Ca de alrededor de 0,6 mg por fruto en la madurez, mientras que los frutos de alta transpiración alcanzarán una concentración de Ca de alrededor de 1,1 mg por fruto. Esta relación no es lineal, el aumento de casi el doble en el Ca de la fruta está asociado con un aumento de casi 6,4 veces en la transpiración de la fruta (Cline y Hanson 1992). Este proceso es restringido por las altas temperaturas en las etapas de desarrollo del fruto, lo cual es muy significativo si revisamos la gráfica de temperaturas estivales en la séptima región de Chile (Figura 5, Fuente: Centro Pomáceas. Universidad de Talca).
CALCIO-ESTRÉS
El ácido abscísico (ABA) se une a un receptor de membrana de las células oclusivas que cierra los estomas a través de la entrada de Ca++ al citoplasma. La síntesis de ABA se eleva y los estomas se cierran hasta que la planta encuentre su equilibrio hídrico (mecanismo de regulación ante estrés hídrico).
Las plantas deben reconocer y responder a las tensiones abióticas, lo que involucra el calcio intracelular. Los cambios inducidos por el estrés en la concentración citosólica de Ca2+ ([Ca2+]cyt) ocurren como resultado de entrada de Ca2+ desde el exterior de la célula o la liberación de Ca2+ de los depósitos intracelulares. Estas alteraciones en [Ca2+]cyt constituyen una señal que se transduce a través de calmodulina, para el ajuste a las nuevas condiciones ambientales.
La señalización de Ca2+ se ha implicado en las respuestas de las plantas a una serie de estrés abióticos ya que los iones de calcio juegan un papel importante como mensajero en el acoplamiento de respuestas fisiológicas (Reddy y Reddy, 2004), cambio en concentración de iones de calcio (calcio libre citosólico). (Rudd y Franklin-Tong, 2001; Sanders et al., 2002).
Kolupaev et al (2005) informaron que la aplicación exógena de Ca2+ promueve la tolerancia al calor de las plantas. Se ha demostrado que la aplicación de calcio en forma de CaCl2 previo al estrés aumenta el contenido de malondialdehído (MDA) (producto de la peroxidación lipídica) y estimula las actividades de guayacol peroxidasa, SOD y catalasa, que podrían ser las razones de la inducción de tolerancia al calor, mediante acción antioxidante. Así mismo bajo estrés por calor, el requerimiento de Ca2+ para el crecimiento es alto para mitigar los efectos adversos del estrés ambiental (Kleinhenz y Palta, 2002), (Waraich et al, 2011). El estrés abiótico conduce a un aumento de calcio libre en el citoplasma de las células.
Revisiones de calcio a cosecha serían indispensables para graficar los niveles de estrés. Altas concentraciones de Ca en los frutos probablemente no determinarán un buen tenor de calcio real, pues podrían inducirnos a un error al relacionarse al calcio proveniente de episodios de estrés vía señalización (probablemente libre).
La determinación de formas del calcio consiste en un proceso de extracción secuencial de cuatro fracciones de calcio de insolubilidad creciente: calcio soluble o libre, calcio ligado (formando pectatos y carbonatos principalmente), calcio insoluble inorgánico (formando fosfatos fundamentalmente) y el calcio insoluble orgánico (en forma de oxalato y silicatos) (Minamide y col., 1986; Carpena y col., 1973; Valenzuela y col., 1994).

Ensayos recientes demuestran que la dificultad de movimiento del calcio hacia el fruto por el xilema que genera la pudrición apical, podría ser una consecuencia primaria de un problema anatómico, ya que puede verse afectada la distribución del calcio dentro del fruto dañado; frutos sanos tienen más calcio soluble que frutos que presentan este desorden fisiológico (Arias, 2018).
El Ca aplicado directamente a la fruta en cerezos fue absorbido y permaneció mayormente dentro de este tejido, sin movimiento hacia dardos. Las observaciones mostraron que los estomas estaban presentes en la superficie inferior de las hojas y, en menor cantidad, en los frutos. Por lo tanto, la presencia de estomas en los órganos es importante para la absorción de Ca, pero este elemento tuvo muy poco movimiento entre los órganos (Bonomelli, 2022).
Es necesario llevar una revisión de las cuatro fracciones de calcio y correlacionarlo con los niveles de estrés de la planta bajo circunstancias controladas y resolver los impactos de estrés en asociación a los tipos de calcio presentes en el fruto, asociado a sus calidades y cualidades.


